Capítulo 6 – DELINEAMENTOS GENÉTICOS
Exercícios:
1) Considere um ensaio de avaliação de g famílias de meios-irmãos, em experimento em blocos ao acaso com r repetições. A parcela é representada por n indivíduos. O esquema da análise de variância é apresentado a seguir:
|
FV |
GL |
QM |
E(QM) |
|
Blocos |
r-1 |
QMB |
|
|
|
|
|
|
|
Famílias |
g-1 |
QMG |
|
|
Entre parcelas |
(r-1)(g-1) |
QME |
|
|
Dentro das parcelas |
(n-1)gr |
QMD |
|
a) Apresente os estimadores dos componentes de variância:
![]()
![]()
![]()
![]()
b) É possível estimar a variância atribuída aos desvios da dominância ?
Não, nesse caso estamos analisando famílias de meios irmãos, onde só é conhecida a porção da variância aditiva.
c) Defina a herdabilidade e apresente um estimador para este parâmetro.
Herdabilidade é dada pela relação entre a variância genética e fenotípica, e representa a porcentagem da variação que é devido a causas genéticas, passiveis de transmissão.
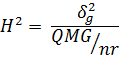
2) Estime, se possível, a variância aditiva e aquela atribuída aos desvios da dominância a partir das seguintes informações:
a) Avaliação de famílias de meios irmãos:
|
FV |
GL |
QM |
E(QM) |
|
Blocos |
2 |
30,42 |
σ² + gσ²b
|
|
Famílias |
99 |
134,67 |
σ² + rσ²g
|
|
Resíduos |
198 |
26,79 |
σ² |
Número de repetições: r= 3; número de famílias: f = 100.
![]()
![]()
![]()
![]()
Não é possível avaliar a variância devida aos desvios de dominância.
b) Avaliação de famílias de irmãos completos:
|
FV |
GL |
QM |
E(QM) |
|
Blocos |
3 |
85,44 |
σ² + gσ²b
|
|
Famílias |
99 |
256,56 |
σ² + rσ²g
|
|
Resíduos |
297 |
101,02 |
σ² |
Número de repetições: r = 4; número de famílias: f = 100
![]()
Considerando σ²d = 0:
![]()
![]()
![]()
c) Avaliação de grupos de machos acasalados com diferentes grupos de fêmeas (hierárquico)
|
FV |
GL |
QM |
|
Machos |
9 |
546,82
|
|
Fêmeas/machos |
50 |
320,60
|
|
Resíduo |
180 |
123,02 |
Número de machos: m = 10; número de fêmeas: f = 6; número de descendentes: n = 4.
![]()
![]()
![]()
i = macho
j = fêmea
k = indivíduo
OBS = utiliza-se a Cov para determinar quais aqueles componentes que estão relacionados, já que a Cov mede a variação comum entre dois termos.
Conhecimento estatístico:
Cov (MI) = E [Mi, M/Fij,
Eijk] [Mi, M/Fij’, Eij’k’]
Cov (MI) = E[Mi²]
Cov (MI) = σ²m
Cov (IC) = E [Mi,
M/Fij, Eijk] [Mi, M/Fij, Eijk’]
Cov (IC) = E[Mi² , M/Fij²]
Cov (IC) = σ²m + σ²f/m
Conhecimento genético:
Cov (MI) = 1/4 σ²a
Cov (IC) = 1/2 σ²a + 1/4 σ²d
Concluindo:
Cov (MI) = 1/4 σ²a = σ²m
σ²a = 4 σ²m
Cov (IC) = 1/2 σ²a + 1/4 σ²d = σ²m + σ²f/m
= 2 σ²m + 1/4 σ²d = σ²m + σ²f/m
= 1/4 σ²d = - σ²m + σ²f/m
σ²d = 4 (σ²f/m - σ²m)
![]()
![]()
![]()
![]()
![]()
![]()
![]()
d) Avaliação de grupos de machos acasalados com um mesmo grupos de fêmeas (fatorial)
|
FV |
GL |
QM |
|
Machos |
9 |
340,84
|
|
Fêmeas |
5 |
410,56
|
|
Machos x Fêmeas |
45 |
120,04
|
|
Resíduos |
180 |
64,32 |
Número de machos: m = 10; número de fêmeas: f = 6; número de descendentes: n = 4.
![]()
![]()
![]()
Conhecimento estatístico:
Cov (IC) = E [Mi,
Fj, MFij, Eijk’] [Mi, Fj, MFij, Eijk’]
Cov (IC) = E[Mi², Fj², MFij²]
Cov (IC) = σ²m + σ²f + σ²mf
Cov (MI) = E [Mi, Fj,
MFij, Eijk’] [Mi, Fj’, MFij’, Eijk’]
Cov (MI) = E[Mi² ]
Cov (MI) = σ²m ou σ²f
Conhecimento genético:
Cov (MI) = 1/4 σ²a = σ²m ou σ²f
σ²a = 2 (σ²m + σ²f) (média dos dois componentes)
Cov (IC) = σ²m + σ²f + σ²mf = 1/2 σ²a + 1/4 σ²d
= 1/4 σ²a
+ 1/4 σ²a + σ²mf = 1/2
σ²a
+ 1/4 σ²d
σ²d = 4 (σ²f/m)
![]()
![]()
![]()
![]()
![]()
![]()
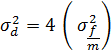
![]()
![]()
e) Avaliação de combinações híbridas de um grupo de genitores.
|
FV |
GL |
QM |
|
CGC |
9 |
544,00
|
|
CEC |
35 |
300,60
|
|
Resíduo |
135 |
120,01 |
Número de genitores: p = 10; número de repetições: r = 4
![]()
![]()
![]()
Modelo = m + Gi + Gj + Sij + Eijk
Conhecimento estatístico:
Cov (IC) = E [Gi, Gj,
Sij, Eijk’] [Gi, Gj, Sij, Eijk’]
Cov (IC) = E[Gi², Gj², Sij²]
Cov (IC) = 2σ²g + σ²s
Cov (MI)
= E [Gi, Gj, Sij, Eijk’] [Gi, Gj’, Sij’, Eij’k’]
Cov (MI) = E[Gi²]
Cov (MI) = σ²g
Conhecimento genético:
Cov (MI) = 1/4 σ²a = σ²g
σ²a = 4 σ²g
Cov (IC) = 1/2 σ²a + 1/4 σ²d = 2σ²g + σ²s
=
1/2 σ²a + 1/4 σ²d = 2(1/4
σ²a) + σ²s
σ²d = 4 σ²s
![]()
![]()
σ²a = 4 σ²g σ²d = 4 σ²s
σ²a = 30,425 σ²d = 180,59
f) Avaliação de gerações
|
FV |
GL |
Média |
Variância |
|
P1 |
50 |
80 |
45 |
|
P2 |
50 |
20 |
56 |
|
F1 |
60 |
70 |
72 |
|
F2 |
400 |
66 |
230 |
|
RC1 |
200 |
72 |
150 |
|
RC2 |
200 |
34 |
180 |
![]()
V (F) =
V(G) + V(M) V (M) =![]()
230 = V(G) + 61,25 V
(M) = ![]()
V(G) = 168,75 V (M) = 61,25
![]()
3) Considere a avaliação de g famílias de meios irmãos, em blocos ao acaso (com r repetições), sendo avaliados n indivíduos dentro da parcela. O esquema da análise de variância é apresentado a seguir:
|
FV |
GL |
QM |
E(QM) |
|
Blocos |
r-1 |
QMB |
|
|
|
|
|
|
|
Famílias |
g-1 |
QMG |
σd² + rnσ²g+ nσ²e
|
|
Entre parcelas |
(r-1)(g-1) |
QME |
σd² + nσ²e
|
|
Dentro das parcelas |
(n-1)gr |
QMD |
σd² |
Forneça
os valores de ( admita ![]() ):
):
![]() Variância fenotípica entre unidades de seleção.
Variância fenotípica entre unidades de seleção.
![]() Variância genética aditiva entre unidade de seleção.
Variância genética aditiva entre unidade de seleção.
|
|
|
FMI |
FIC |
||
|
Estratégia |
Unid. Seleção |
δ²g |
δ²f |
δ²g |
δ²f |
|
Entre famílias |
Yi.. x Yi’.. |
¼ δ²a (δ²g) |
QMG/nr |
½ δ²a |
QMG/nr |
|
Dentro família |
Yijk x Yijk’ |
¾ δ²a (3 δ²g) |
QMD |
½ δ²a |
QMD |
|
Massal |
Yijk x Yi’j’k’ |
4 δ²g |
δ²g+ δ²b+ δ²e+ δ²d |
2 δ²g |
δ²g+ δ²b+ δ²e+ δ²d |
|
Massal estratificada |
Yijk x Yi’jk’ |
4 δ²g |
δ²g + δ²e+ δ²d |
2 δ²g |
δ²g + δ²e+ δ²d |
4) Considere a avaliação de g famílias de irmãos completos em blocos ao acaso (com r repetições), sendo avaliados n indivíduos dentro da parcela. O esquema da análise de variância é apresentado a seguir:
|
FV |
GL |
QM |
E(QM) |
|
Blocos |
1 |
12 |
|
|
Famílias |
19 |
60 |
|
|
|
|
|
|
|
Entre parcelas |
19 |
10 |
|
|
Dentro das parcelas |
80 |
5 |
|
Admita que a variância atribuída aos desvios da dominância seja nula e forneça os valores de:
![]() Variância fenotípica entre as unidades de seleção.
Variância fenotípica entre as unidades de seleção.
![]() Variância genética aditiva entre as unidade de seleção.
Variância genética aditiva entre as unidade de seleção.
r = 2
g =20
(n-1)gr = 80
(n-1) 40 = 80
n = 3
![]()
![]()
![]()
![]()
|
Estratégia |
Unid. Seleção |
δ²g |
δ²f |
|
Entre famílias |
Yi.. x Yi’.. |
δ²g = 8,33 |
QMG/nr = 10 |
|
Dentro família |
Yijk x Yijk’ |
δ²g = 8,33 |
QMD = 5 |
|
Massal |
Yijk x Yi’j’k’ |
2 δ²g = 16,66 |
δ²g+ δ²b+ δ²e+ δ²d = 15,0333 |
|
Massal estratificada |
Yijk x Yi’jk’ |
2 δ²g = 16,66 |
δ²g + δ²e+ δ²d = 15 |
5) Considere as informações apresentadas a seguir, obtidas da avaliação de grupos de machos acasalados com diferentes grupos de fêmeas.
|
FV |
GL |
QM |
|
Machos |
9 (m-1) |
400 |
|
Fêmeas/machos |
50 (f-1)m |
200 |
|
Resíduos |
180 (n-1)mf |
100 |
|
|
|
|
Número de machos: m=10; número de fêmeas: f=6; número de descendentes: n=4.
Estime:
a) Herdabilidade no sentido amplo para a seleção de machos.
Neste caso, a seleção é praticada entre os valores médios obtidos para cada macho, comparando-se Yi.. e Yi’.., que podem ser expressos por meio de:
Yi.. = m
+ Mi + 1/f Σ F/Mij + 1/nf ΣΣEijk
Yi’.. = m
+ Mi’ + 1/f Σ F/Mi’j + 1/nf ΣΣEi’jk
Δ (Var. fenotípica) = δ²f + δ²f/m /f + δ²d /nf = QMM/nf
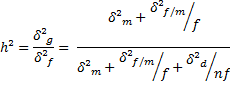
![]()
![]()
![]()
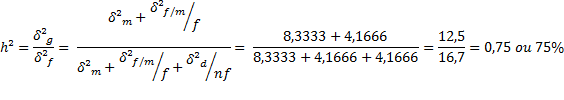
b) Variância genética aditiva.
Conhecimento estatístico:
Cov (MI) = E [Mi, M/Fij,
Eijk] [Mi, M/Fij’, Eij’k’]
Cov (MI) = E[Mi²]
Cov (MI) = σ²m
Cov (IC) = E [Mi,
M/Fij, Eijk] [Mi, M/Fij’, Eij’k’]
Cov (IC) = E[Mi² , M/Fij²]
Cov (IC) = σ²m + σ²f/m
Conhecimento genético:
Cov (MI) = 1/4 σ²a
Cov (IC) = 1/2 σ²a + 1/4 σ²d
Concluindo:
Cov (MI) = 1/4 σ²a = σ²m
Cov (IC) = 1/2 σ²a + 1/4 σ²d = σ²m + σ²f/m
σ²a = 4 σ²m
σ²a = 4 (8,3333)
σ²a = 33,33
c) Variância atribuída aos desvios de dominância.
σ²d = 4 (σ²f/m - σ²m)
σ²d = 4 (25-8,3333)
σ²d = 66,67
6) Estime a herdabilidade no sentido amplo e restrito, para a seleção entre médias de machos, considerando as informações apresentadas a seguir, obtidas da avaliação de grupos de machos acasalados com diferentes grupos de fêmeas.
|
FV |
GL |
QM |
|
Machos |
9 (m-1) |
380,50
|
|
Fêmeas/machos |
50 (f-1)m |
120,60
|
|
Resíduo |
180 (n-1)mf |
62,40 |
|
|
|
|
Número de machos: m=10; número de fêmeas: f=6; número de descendentes: n=4.
Neste caso, a seleção é praticada entre os valores médios obtidos para cada macho, comparando-se Yi.. e Yi’.., que podem ser expressos por meio de:
Yi.. = m
+ Mi + 1/f Σ F/Mij + 1/nf ΣΣEijk
Yi’.. = m
+ Mi’ + 1/f Σ F/Mi’j + 1/nf ΣΣEi’jk
Δ (Var. fenotípica) = δ²f + δ²f/m /f + δ²d /nf
![]()
![]()
![]()
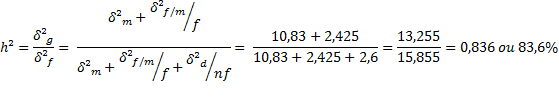
Conhecimento estatístico:
Cov (MI) = E [Mi, M/Fij,
Eijk] [Mi, M/Fij’, Eij’k’]
Cov (MI) = E[Mi²]
Cov (MI) = σ²m
Cov (IC) = E [Mi,
M/Fij, Eijk] [Mi, M/Fij’, Eij’k’]
Cov (IC) = E[Mi² , M/Fij²]
Cov (IC) = σ²m + σ²f/m
Conhecimento genético:
Cov (MI) = 1/4 σ²a
Cov (IC) = 1/2 σ²a + 1/4 σ²d
Concluindo:
Cov (MI) = 1/4 σ²a = σ²m
Cov (IC) = 1/2 σ²a + 1/4 σ²d = σ²m + σ²f/m
σ²a = 4 σ²m
σ²a = 4 (10,83)
σ²a = 43,32
σ²d = 4 (σ²f/m - σ²m)
σ²d = 4 (14,55 – 10,83)
σ²d = 14,88
Herdabilidade Restrita:
![]()
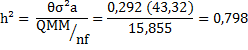
7) Quando são feitas análises de variâncias, obtêm-se informações sobre vários parâmetros de uma população de interesse. Qual a utilidade dos parâmetros abaixo para você (na qualidade de melhorista)?
a) Média da população
O ganho de seleção, predito nos programas de melhoramento é função da herdabilidade e do diferencial de seleção. A herdabilidade é um parâmetro dependente da característica em estudo, já o diferencial de seleção é dado pela diferença entre a média total da população e a média das populações selecionadas pelo melhorista. Dessa forma, o conhecimento da média é importante para a seleção das melhores populações ou progênies, o que resulta em maiores ganhos de seleção para o programa.
b) Coeficiente de variação experimental
Pode ser calculado dividindo a raiz quadrada do QMR pela média, indicando assim a precisão do experimental. Quanto menor o valor de CV melhor o experimento.
c) Variância genética e valor da estatística F
A variância genética é aquela porção da nossa variância total (fenotípica) que é herdada. Ela ainda pode ser subdivida na sua parte aditiva e também naquela causada pelos desvios de dominância. Seu conhecimento permite que o melhorista calcule a herdabilidade de um caráter e, consequentemente, estime o ganho de seleção.
O valor da estatística F visa testar valores de variância obtida, sendo importante nas tomadas de decisões. Valor significativo indica rejeição da hipótese inicial, enquanto valor não significativo indica aceitação desta hipótese.
d) Herdabilidade
Razão entre a variância genética e fenotípica, indica ao melhorista a porcentagem realmente herdável de uma característica. Muito importante para o calculo de ganho de seleção, sendo que, quanto maior o valor de herdabilidade, maior o ganho de seleção.
8) Uma das grandes contribuições da Genética Quantitativa é a predição de ganhos. Uma população apresenta média igual a 80, porém os estudos genéticos indicam que, ao aplicar determinada técnica de seleção, o ganho será de 8 e, consequentemente, a média da população melhorada será de 88. Você acredita que, de fato, ao se obter e avaliar uma população melhorada, ela produzirá os 88 preditos? Qual a utilidade da predição de ganho a ser obtido pelo melhorista?
Não. Dificilmente o melhorista observa na pratica aquilo que foi previsto na teoria, uma vez que, durante a formulação teórica dos métodos quantitativos alguns fatores genéticos são negligenciados, sendo sempre o valor observado diferente do esperado. No entanto a predição de ganho é importante para o melhorista, pois é um indicativo do que esperar, além de permitir que o profissional escolha o melhor método de melhoramento.
9) Recentemente tem sido considerado que grandes avanços na área de melhoramento serão proporcionados por estudos de Genética Molecular. Você julga serem dispensáveis os conhecimentos sobre análise biométrica para os profissionais desta área? Justifique.
Não. A genética molecular e biometria devem andar atrelados auxiliando os programas de melhoramento. Informações moleculares são importante, pois acessam a parte do genoma que é realmente herdável, sem a ação de agentes perturbadores como o meio ambiente, por exemplo. No entanto, essas informações são úteis apenas se puderem ser aplicadas diretamente no melhoramento convencional, que é responsável pelo desenvolvimento e lançamento de cultivares. E para realização do melhoramento convencional, é importante que o melhorista tenha conhecimento de biometria para maximizar os ganhos.
10) Os dados do quadro a seguir foram obtidos em um programa de melhoramento de milho-pipoca e referem-se a produção de grãos (PG) e à capacidade de expansão (CE) de famílias de meios irmãos.
|
FV |
GL |
QMPG |
QMCE |
|
Blocos |
3 |
- |
- |
|
FMI |
99 |
13172,3 |
41,71 |
|
Resíduo |
297 |
6262,68 |
19,08 |
|
Média
|
|
179,8 (kg/ha) |
24,9 (ml/g) |
a) Quais as estimativas da variância genotípica entre progênies para as duas características?
Para PG:
![]()
Para CE:
![]()
b) Estime as herdabilidades das características.
Para PG:
![]()
![]()
Para CE:
![]()
![]()