GENÉTICA MOLECULAR
|
|
·
Produção: Laboratório de Bioinformática
·
Aplicativo suporte: Programa GBOL – Genética Básica on
line
·
Comunidade (facebook): GbolNews
|

Tópicos
Identificação do material genético
DNA e RNA
Função do material genético
Bases fisiológicas da dominância e recessividade
Código genético
Síntese de cadeia polipeptídica
IDENTIFICAÇÃO DO MATERIAL GENÉTICO
Em 1866,
Mendel descreveu os genes pelos seus efeitos finais, tais como os
fenótipos. Pelos experimentos de Mendel e de outros pesquisadores, ficou
definido que os genes portam informação genética e, apesar de não serem vistos ou
delimitados fisicamente, deveriam apresentar as seguintes propriedades:
- Replicação: processo que permite ao gene produzir outras unidades iguais a si próprio.
- Transcrição: processo pelo qual a informação genética é transferida para o
local apropriado (ribossomo) e é traduzida.
-
Tradução: processo pelo qual são produzidas as proteínas, a partir de uma seqüência de nucleotídeos.
Após Mendel, os genes foram definidos quimicamente e conhecidos pelo que
realizam na síntese protéica e não pelo nível de
expressão fenotípica.
Miescher (1869-71) publicou uma metodologia que permite separar o
núcleo do citoplasma. Do núcleo, ele extraiu uma substância denominada nucleína, hoje conhecida por ácido nucléico, que se
caracterizava por apresentar alta acidez e grande quantidade de fósforo e não
continha enxofre.
Mais tarde, descobriram que a nucleína estava
associada a vários tipos de proteínas, formando a nucleoproteínas.
Da porção protéica foram constatados dois tipos de
proteínas:
a. Protaminas: proteínas de estrutura simples,
constituídas, na maioria das vezes, de
grupos do aminoácido arginina. Essa proteína está presente no esperma de peixes
e aves.
b. Histona: proteínas relativamente complexas de
ocorrência mais ampla.
Embora os peixes e as aves não sejam as mais complexas das criaturas, parece
difícil aceitar a idéia de que o material genético
desses organismos seja a protamina e que, constituída
quase que apenas de arginina, ela seja capaz de originar os 20 aminoácidos
conhecidos. Também é pouco aceitável que o material genético seja a histona,
até a formação de uma protamina.
Griffith
(1928) apresentou as primeiras evidências de que o DNA é o material genético.
As evidências surgiram em experimentos realizados com bactérias do gênero Pneumococus. Muitas linhagens ou tipos de pneumococus (Diplococcus
pneumoniae) podem ser distinguidos por diversas
características:
|
Caracterização |
Virulenta |
Avirulenta |
|
Colônia |
Lisa (S) |
Rugosa (R) |
|
Capa protéica |
Presente |
Presente |
O experimento realizado por Griffith é resumido a
seguir:
|
Tratamento |
Resultado |
|
· Tipo II R injetado em ratos |
ratos sobreviveram |
|
· Tipo III S morto pelo calor injetado em rato |
ratos sobreviveram |
|
· Tipo III S morto pelo calor mais o tipo II R
injetado em ratos |
Ratos morreram |
A mudança não poderia ter surgido por mutação, pois um mutante deveria ser da
mesma linhagem genética do tipo III, entretanto foi o tipo II (com capa protéica) que adquiriu a propriedade de causar a doença tal
como o tipo III. Algum fator das bactérias do tipo III S estava aparentemente
sendo transferido para os cocus vivos (tipo II), de forma que o tipo II avirulento transformara-se em cocus virulentos. Esse
fenômeno foi conhecido como "efeito Griffith" ou
"transformação".
Avery, MacLeod e McCarty (1944) publicaram resultados de extensas
investigações durante um período de 10 anos. Eles fizeram, em condições de
laboratório, experimentos semelhantes ao de Griffith. Avery
e seus colaboradores relataram que ao colocar em um tubo de ensaio bactérias II
R (avirulenta), extrato de DNA do tipo III S
(virulenta) mortas pelo calor e soro que precipitava as bactérias do tipo II R,
foram recuperadas as colônias de bactérias do tipo III S. Esses experimentos
identificaram o DNA como material genético. Assim, quando o DNA extraído de uma
linhagem foi introduzido em células de outra linhagem, os organismos receptores
desenvolveram características da linhagem doadora. Dessa forma, conclui-se que:
- O DNA
é o material genético em D. pneumoniae.
- O DNA atua como agente que especifica produção de um produto final.
![]() Volta
Volta
DNA E RNA
Constituição química do DNA
e RNA
Quando o ácido nucléico foi separado da proteína, muitos pesquisadores,
especialmente Levene, mostraram que ele poderia ser
quebrado em pequenas partes denominadas nucleotídeos. Cada nucleotídeo contém:
|
Caracterização |
DNA |
RNA |
|
1. Açúcar |
Desoxirribose |
Ribose |
|
2. Grupo Fosfato |
H3PO4 |
H3PO4 |
|
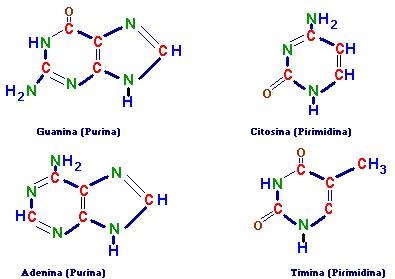
3. Bases nitrogenadas |
Pirimidinas: |
Pirimidinas: |
Chargaff, em seus estudos, apresentou várias
informações a respeito da molécula de DNA. Mostrou que ocorriam ligações entre
a pirimidina citosina e purina guanina e entre a pirimidina timina e a purina
adenina. As ligações envolvem duas pontes entre A e T e três entre G e C.
Também constatou a seguinte relação:
(C+A)/(G+T) =1
Diferenças entre DNA e RNA
O DNA se
diferencia do RNA nos seguintes aspectos:
a. O
açúcar do DNA é a desoxirribose, enquanto
o do RNA é a ribose.
b. O DNA
contém a timina e o RNA a uracil.
c. O DNA
é um filamento duplo e o RNA é um monofilamento.
d. O DNA
apresenta uma molécula longa e o RNA, uma molécula curta.
Modelo do DNA segundo Watson
e Crick
Watson e Crick propuseram um modelo de DNA cujos princípios físicos derivaram
das figuras de difração de raios X produzidas por Wilkins e Astracan,
usando DNA isolado. Os princípios químicos derivaram principalmente dos
trabalhos apresentados por Chargaff e seus associados.
Segundo Watson e Crick, o DNA apresenta as seguintes características:

a. O DNA é uma hélice dupla helicoidal.
b. O
enrolamento da hélice é para a direita.
c. Os
longos filamentos externos, relativamente rígidos, são constituídos de fósforo
(P) e açúcar (A).
d. No
sentido transversal, os filamentos menos rígidos são constituídos por bases
orgânicas (purinas e pirimidinas) unidas por pontes de hidrogênio.
e. O comprimento de uma volta completa na espiral
envolve cerca de 10 nucleotídeos (34 angstron).
Associação entre DNA e
Histonas
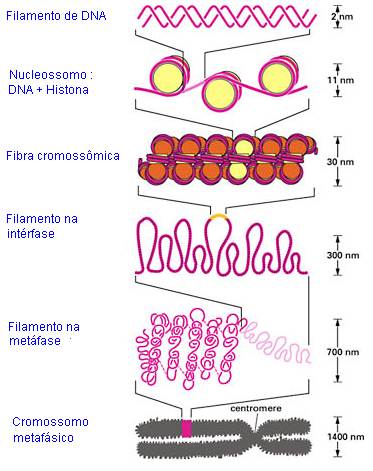
As histonas formam um complexo juntamente com os grupos fosfatados do DNA carregados negativamente. As histonas são carregadas positivamente, sendo conhecidas por "proteínas básicas". As cargas positivas são fornecidas por alta proporção de aminoácidos lisina e arginina. Algumas histonas são denominadas "ricas em lisina" e outras "ricas em arginina".
Em geral são encontradas somente nos organismos em que a diferenciação celular
ocorre (eucariotas). São distinguidos, em função da proporção lisina/arginina,
cinco diferentes tipos de histonas (H1, 2 H2A, 2 H2B e
2 H3).
A complexação das histonas, além de causar um aumento
do diâmetro do DNA, de cerca de 20 a 30 angstron,
muda também suas propriedades. A temperatura de fusão (temperatura na qual os fios
de DNA mudam da forma de hélice dupla regular para a forma de fio simples) é
bastante aumentada.
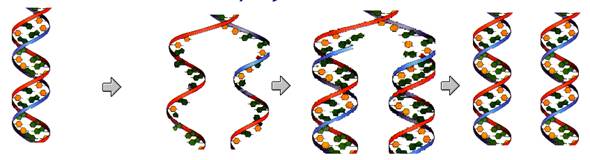
Replicação do material
genético
O modelo
de replicação do DNA é dito ser semiconservativo, ou seja, uma fita de DNA dá
origem a outras duas em que cada uma delas apresenta um filamento da fita
original.
A replicação ocorre na intérfase da divisão celular e
é dirigida pela enzima DNA polimerase. Na replicação, as pontes de hidrogênio
entre as bases nitrogenadas se rompem e a enzima DNA polimerase catalisa a
adição de novos nucleotídeos complementares às bases expostas, de maneira que
duas novas fitas de DNA sejam formadas. .
O RNA - Ácido ribonucléico
O RNA é
encontrado tanto no núcleo quanto no citoplasma. Têm sido reconhecidos vários
tipos de RNA:
a. RNA mensageiro - mRNA
É o RNA
envolvido pela transcrição da informação genética contida no DNA, no núcleo e
condução da mesma até os sítios ribossômicos.
b. RNA transportador - tRNA
Caracteriza-se
por ser uma molécula pequena, contendo cerca de 75 a
85 nucleotídeos e por assumir forma de trevo. Sua função principal é a de
conduzir os aminoácidos requeridos na síntese protéica
até as subunidades do ribossomo.
c. RNA ribossômico - rRNA
Participa na composição do
ribossomo. Sua função não é bem definida.
![]() Volta
Volta
FUNÇÃO DO MATERIAL GENÉTICO
Garrod (1900), médico inglês, deu os primeiros passos
para evidenciar a ação dos genes. Esse pesquisador estudou a alcaptonúria, doença hereditária caracterizada pela
coloração escura na urina. No caso dessa doença, foi observado que o
escurecimento era conseqüência do acúmulo de ácido homozentízico (alcapton), o qual,
nos indivíduos normais, era decomposto através de reações enzimáticas, havendo
excreção de ácido acético. Constatou-se ainda que dietas com tirosina e fenilalamina aumentava a
manifestação de sintomas.
Os
trabalhos decisivos sobre a função do gene foram apresentados por Beadle e
Tatum, que propuseram a teoria "um gene - uma enzima". As idéias principais do trabalho realizado por esses autores
são:
a.
Todos os
processos bioquímicos dos organismos estão sob controle
genético.
b. Os processos bioquímicos ocorrem numa seqüência de reações individuais.
c. Cada reação simples é controlada por um gene simples.
d.
Cada gene atua
por meio do controle e produção de uma enzima específica.
Na atualidade essa teoria apresenta as seguintes falhas:
a. Um
gene pode especificar a síntese de uma cadeia polipeptídica que não apresenta
nenhuma função enzimática (Ex. hemoglobina).
b. Uma enzima
pode ser constituída por mais de uma cadeia polipeptídica (Ex
RNA polimerase é constituída por várias cadeias e, conseqüentemente,
está sob o controle de vários genes.
c. Um
gene pode controlar a atividade de uma enzima especificada por outro gene (Ex.
sítios operadores, repressores etc.).
![]() Volta
Volta
BASES FISIOLÓGICAS DA DOMINÂNCIA E RECESSIVIDADE
De
acordo com a teoria "um gene - uma enzima", os genes agem através da
produção de enzimas, sendo
cada gene responsável pela
produção de uma enzima específica. Por esse princípio, pode-se explicar as
bases fisiológicas da dominância e recessividade, segundo determinada via biossintética. Assim,
tem-se:
Dominância completa
A
quantidade de enzimas produzidas pelo heterozigoto Aa,
embora geralmente menor que a produzida pelo homozigoto AA, é suficiente para
produção do mesmo produto final de AA.
Dominância. incompleta
A menor quantidade de
enzimas normal produzida pelo heterozigoto Aa, em relação ao homozigoto AA,
resulta em menor quantidade de produto
final que, pelo efeito de dosagem, confere um fenótipo diferente do produzido
pelo homozigoto AA.
Codominância
O complexo enzimático produzido por Aa, dadas as
contribuições de A e de a, é diferente da enzima produzida por AA, e, conseqüentemente o produto final será diferente daquele
produzido por AA. Então, a qualidade de enzima é o fator crítico.
![]() Volta
Volta
CÓDIGO GENÉTICO
Definição de código genético
Sabe-se
que o DNA, que se encontra no núcleo, tem a função de produzir proteínas cuja
síntese ocorre no citoplasma. No DNA, que é uma seqüência
de nucleotídeos, existem
apenas quatro tipos diferentes: os nucleotídeos da adenina, da guanina, de
timina e da citosina. No entanto, as proteínas são polímeros de
subunidades (monômeros) denominadas
aminoácidos. Cada aminoácido engloba um grupo amino (NH2)
numa extremidade e um grupo carboxila (COOH) na outra. Vinte tipos diferentes
de aminoácidos ocorrem nas proteínas.
Diante do exposto, surge a seguinte pergunta: quantos nucleotídeos seriam necessários
para codificar um aminoácido? Para responder a essa pergunta, é necessário o
seguinte raciocínio matemático:
- Se 1 nucleotídeo codificasse um aminoácido, só
poderiam existir 4 diferentes tipos de aminoácidos na cadeia protéica.
- Se 2 nucleotídeos codificassem um aminoácido, só poderiam
existir 16 tipos de aminoácidos diferentes na cadeia protéica.
- Se 3 aminoácidos codificassem um aminoácido, seria possível
haver 64 tipos diferentes de aminoácidos na cadeia protéica;
logo, por matemática, um código tríplice é a menor unidade de codificação capaz
de acomodar os 20 diferentes tipos de aminoácidos que comumente ocorrem nas
proteínas.
Atualmente
define-se códon
como uma seqüência
de três nucleotídeos adjacentes no mRNA capaz de
codificar um aminoácido.
Decifração do código
genético
Estudos permitiram constatar que o código genético é representado por 64 códons mRNA, 61 dos quais
codificam para aminoácidos e três indicam o
término da síntese protéica. Três dos que
codificam aminoácidos são identificados também como iniciadores.
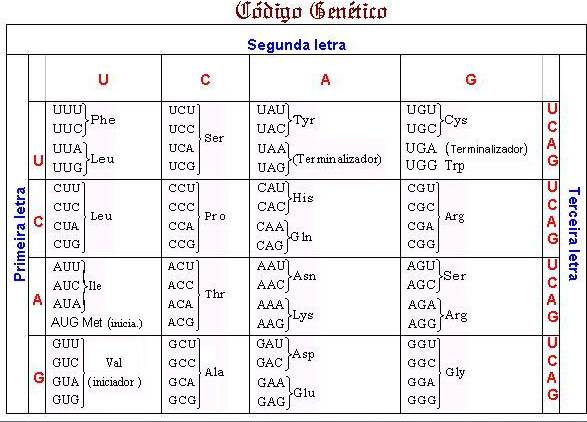
São evidenciadas as seguintes características do código genético:
Código genético redundante ou degenerado
O código
genético é dito degenerado pelo fato de existir, para um determinado
aminoácido, mais de uma trinca que o codifique. Apenas a metionina (Met) e o triptofano (Trp) são codificados por um único códon e representados por
AUG e UGG, respectivamente.
A glicina (GLY), por exemplo, é codificada por GGG,
GGC, GGA e GGU.
Trincas sem sentido ou terminalizadoras
São
aquelas que não
codificam aminoácidos e que têm por função indicar o término da síntese protéica. São também denominados trincas terminalizadoras. Ex.: UAG, UAA, UGA.
Código genético universal
O código
genético é dito universal devido ao fato de a mesma trinca codificar o mesmo
aminoácido, em
qualquer organismo. Em alguns casos, certas trincas são mais eficientemente
utilizadas.
![]() Volta
Volta
SÍNTESE DE CADEIA POLIPEPTÍDICA
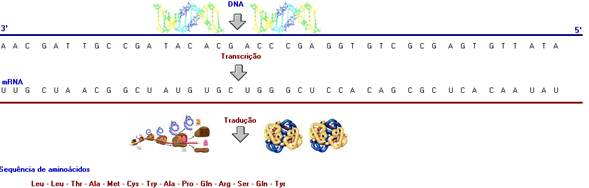
A síntese protéica envolve as seguintes etapas:
Transcrição e Tradução
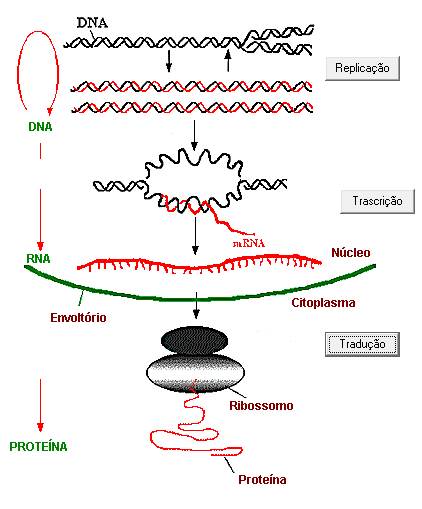
Transcrição
Processo que ocorre no núcleo da célula, no qual a mensagem genética é passada
do DNA para uma fita de mRNA,
com auxílio da ação catalítica da enzima RNA polimerase.
A informação do DNA é transcrita em uma seqüência
codificada do mRNA, através
do uso, com gabarito, de um filamento do DNA. Até hoje, o mecanismo de seleção
do filamento apropriado
é desconhecido.
O mRNA transcrito solta-se
do modelo de DNA e desloca-se do núcleo para o citoplasma. Após o mRNA se desacoplar, as pontes de
hidrogênio que haviam sido desfeitas, voltam a se ligar.
A enzima RNA polimerase tem as seguintes funções:
a.
Reconhecer as bases do DNA.
b.
Selecionar os ribonucleotídeos apropriados.
c.
Catalisar a formação de ligações entre os ribonucleotídeos.
d.
Escolher o filamento correto a ser transcrito.
A RNA polimerase é constituída de seis cadeias polipeptídicas (2 alfa, beta, beta', gama e sigma), sendo o fator sigma
o responsável pela transcrição no local e fio correto.
Tradução
A tradução é um processo que ocorre no citoplasma da célula, no qual a mensagem
trazida pela fita de mRNA é
traduzida em uma seqüência de aminoácidos.
O
processo de tradução envolve as seguintes etapas:
a. Após a chegada da fita de mRNA
no citoplasma, ocorre a complexação das subunidades
do ribossomo (nos procariotas o ribossomo é 70 s = 50 s + 30 s, e nos
eucariotas é de 80 s = 50 s + 40 s) com essa fita. Polissomos é a denominação
que se dá à complexação de vários ribossomos a uma
mesma fita de mRNA. No caso
da síntese da hemoglobina, os ribossomos reúnem-se em quatro ou cinco
polissomos.
b. São iniciadas a leitura e a tradução da fita de mRNA. Nos procariotas, em geral, a primeira trinca a ser
lida (fator de inicialização) é a AUG, que corresponde à metionina formilada (As trincas GUU, GUC, GUA e GUG que correspondem
à valina também são fatores iniciadores), enquanto nos eucariotas o primeiro aminoácido é
também a metionina, mas não formilada. Nem todas as
proteínas se iniciam com a metionina (ou valina), e isso se dá pelo fato de a
proteína final ser o resultado de uma reestruturação, incluindo quebras, da
estrutura linear de aminoácido.
Após a leitura da trinca, ocorre a transferência do aminoácido requerido para
os sítios ribossômicos. Esse transporte é realizado pelo tRNA. No tRNA
é encontrado o anticódon, que é uma seqüência de três
bases adjacentes complementares ao códon do mRNA.
c. O tRNA, penetrando por
uma abertura da subunidade 50s, ocupa o sítio aminoacil.
Pela ação da enzima translocase, o tRNA ativado passa para o sítio peptidil, permitindo a leitura, pelo complexo ribossomo-mRNA, da trinca consecutiva.
d. Após a
leitura da nova trinca, um novo tRNA é ativado e
deslocado do suco citoplasmático para o sítio aminoacil
do complexo, levando o aminoácido especificado
pelo códon.
e. Pela ação da enzima peptidil transferase,
o aminoácido (ou seqüência de aminoácidos) que
pertencia ao tRNA do sítio peptidil é transferido e ligado ao aminoácido acoplado ao tRNA do sítio aminoacil.
f. O tRNA desativado, que ocupa o sítio peptidil,
deixa o complexo ribossomo-mRNA, podendo ser
novamente ativado quando se fizer necessário.
g.
Novamente, pela ação da enzima translocase, o tRNA ativado passa do sítio aminoacil
para o sítio peptidil, permitindo a leitura de uma
nova trinca. A leitura da nova trinca implica ativação de um outro tRNA.
h. O
processo continua até que todos os
aminoácidos necessários para a confecção da proteína estejam ligados. A última
trinca lida na fita de mRNA
deverá ser um fator de terminalização (UAA, UAG ou
UGA), que não codifica nenhum aminoácido mas indica o término da síntese protéica.
i. A síntese protéica
termina com a desativação do complexo ribossomo-mRNA e formação da
proteína que ainda se encontra em forma linear. Para adquirir a especificidade,
é necessário que essa estrutura primária atinja uma estrutura terciária ou
quaternária.