MECANISMOS DE
DETERMINAÇÃO DO SEXO
|
|
·
Produção: Laboratório de Bioinformática
·
Aplicativo suporte: Programa GBOL – Genética Básica on
line
·
Comunidade (facebook): GbolNews
|

Tópicos
Introdução
Mecanismo dos Cromossomos Sexuais
Balanço Gênico
Haplodiploidismo
Efeito de Genes
Efeito do Ambiente
Aberrações Sexuais na Espécie Humana
Cromatina Sexual
Ginandromorfismo
INTRODUÇÃO
O caráter sexo sempre foi tratado como apresentando um tipo especial de
herança, uma vez que em qualquer cruzamento, para a maioria das espécies, o
resultado apresentado pela progênie é sempre 1/2 macho :
1/2 fêmeas. A idéia da existência de vários mecanismos
envolvidos na determinação do sexo resultou de vários trabalhos envolvendo diferentes
espécies vegetais e animais.
Alguns dos trabalhos fundamentais foram realizados por Van Beneden
em 1866, relatando que a fertilização deveria envolver a união de um espermatozóide e um óvulo. Estes resultados foram obtidos
em experimentos realizados em coelhos. Hartwig (1876)
descreveu o mesmo fato em experimentos envolvendo ouriço-do-mar.
Henking (1891) apresentou as primeiras investigações
que relacionaram cromossomos com determinação do sexo. Seus estudos, realizados
com insetos, revelaram que metade dos espermatozóides
recebia determinada estrutura nuclear e a outra metade não. Ele chamou esta estrutura de
corpúsculo X. McClung (1902) realizou estudos
citológicos em diferentes espécies de gafanhotos e concluiu que as células
somáticas das fêmeas tinham número de cromossomos diferente das células do
macho e que havia associação entre a presença do cromossomo X e a determinação
de sexo nesses insetos.
Wilson, trabalhando com insetos do gênero Protenor,
demonstrou que as fêmeas apresentavam 14 cromossomos em suas células somáticas
e eram capazes de produzir gametas carregando 7
cromossomos. Os machos apresentavam 13 cromossomos em suas células somáticas e
eram capazes de produzir gametas com 7 ou 6
cromossomos. A diferença do número de cromossomos era determinante do sexo.
![]() Volta
Volta
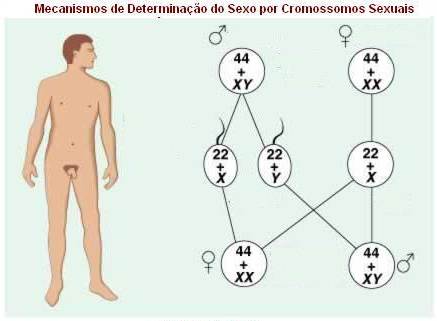
MECANISMO DOS CROMOSSOMOS SEXUAIS
Macho heterogamético -
Espécie humana e outros mamíferos
Na espécie humana, e aparentemente em todos os outros mamíferos, a presença do
cromossomo Y determina a masculinidade. Na espécie humana, o sexo é definido da
seguinte maneira:
|
Sexo |
Célula Somática |
Célula Gamética |
|
Homem |
44 autossomais + XY |
22 autossomais + X ou Y |
|
Mulher |
44 autossomais + XX |
22 autossomais + X |
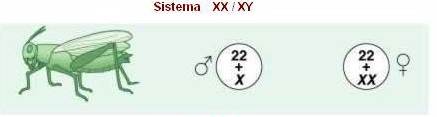
Macho heterogamético - Insetos:
percevejos (Hemípteros),
gafanhotos e baratas (Orthoptera)
Neste caso, o número de cromossomos X é o determinante do sexo. O sexo é
caracterizado por:
Fêmeas:
- apresentam número
par de cromossomos em suas células somáticas;
-são do
tipo XX + 2A; e
-produzem
apenas um tipo de gameta: A + X.
Machos:
- apresentam número ímpar de cromossomos em suas células somáticas;
-são do
tipo: 2A + XO; e
-produzem
dois diferentes gametas: (A + X) e (A).
Também,
neste caso, a proporção ½ macho: ½ fêmea é mantida, pois os cruzamentos
envolvem os indivíduos XX x XO.
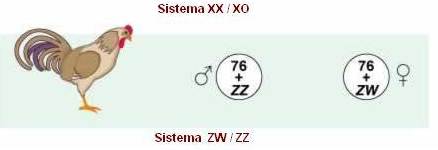
Fêmea heterogamética
- borboletas, mariposas e alguns peixes e aves
Neste
caso, a determinação de sexo se assemelha à apresentada pela espécie humana,
entretanto, neste caso, as fêmeas são heterogaméticas. Por motivo de clareza, o
cromossomo X passa a ser representado por Z, e o Y, por W. Os sexos são
caracterizados por:
Fêmeas
- apresentam constituição cromossômica: 2A + ZW; e
-
produzem dois tipos diferentes de gametas: (A + Z) e (A + W).
Machos
- apresentam constituição cromossômica: 2A + ZZ; e
-
produzem apenas um tipo de gameta:( A + Z).
![]() Volta
Volta
BALANÇO GÊNICO
O mecanismo de determinação de sexo pelo balanço gênico é aplicável aos insetos
do gênero Drosophila. Inicialmente imaginou-se
que o mecanismo de determinação de sexo nesses insetos seria semelhante ao
apresentado pela espécie humana. Em observações citológicas de células somáticas,
constatou-se um conjunto diplóide de 8 cromossomos (2x = 8), em que as fêmeas apresentam
constituição cromossômica 2A + XX, e os machos, 2A + XY.
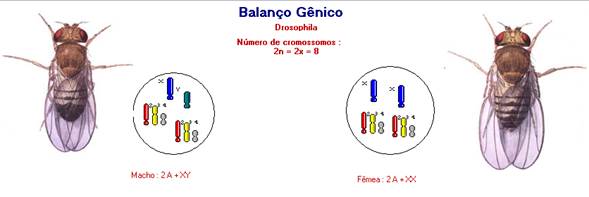
Com base em observações em tipos sexuais, foi proposto que a determinação do
sexo em Drosophila seria função de um índice
sexual (Is), que é função do balanço entre
cromossomos X e conjuntos autossomais, conforme
descrito a seguir:
IS = (Número de cromossomos X)/(Número de conjuntos
autossomais)
Com base nesse índice sexual, o sexo seria determinado segundo a tabela abaixo:
|
Índice Sexual (IS) |
Sexo
|
|
< 0,5 |
Metamacho |
|
0,5 |
Macho |
|
(0,5 - 1,0) |
Intersexo |
|
1,0 |
Fêmea |
|
> 1,0 |
Metafêmea |
![]() Volta
Volta
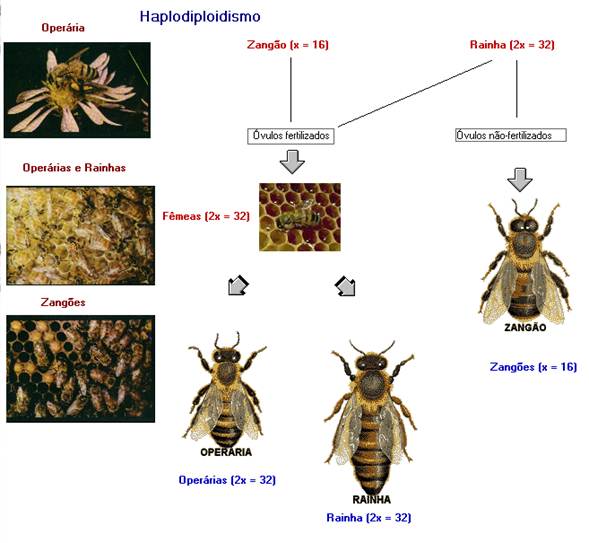
HAPLODIPLOIDISMO
É um complexo mecanismo de determinação de sexo descrito para Himenópteros (formigas,
abelhas e vespas). Nas abelhas, o sexo masculino é definido pela condição haplóide (ou monoplóide) e o sexo
feminino, pela condição diplóide do indivíduo. As
fêmeas, organismos diplóides, dependem da alimentação
para adquirir fertilidade. A alimentação prolongada com geléia
real possibilita a formação de rainhas que são responsáveis pela formação da colméia.
Série de alelos múltiplos em
abelhas
Em adição ao sistema haplodiplóide de determinação do
sexo existe uma série de alelos múltiplos também envolvida
nas manifestações sexuais. A série é representada por 12 alelos: S1,
S2, ...... S12 e atua da
seguinte maneira:
a. Indivíduos resultantes da fertilização
Heterozigotos (SiSj, para i diferente de j)
serão fêmeas
Homozigotos (SiSj para i = j) serão machos
inviáveis.
b.Indivíduos não fertilizados: Serão machos hemizigóticos.
![]() Volta
Volta
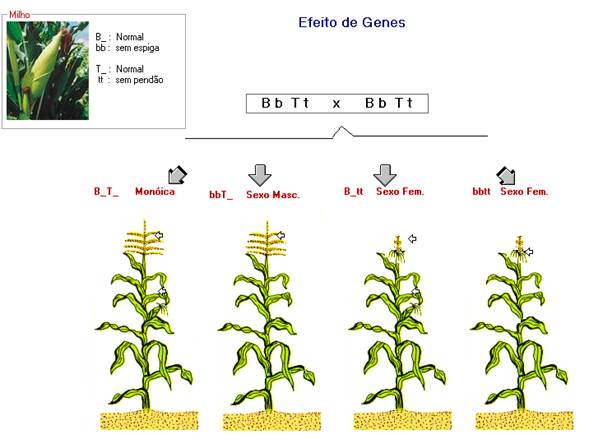
EFEITO DE GENES
Neste caso, a determinação do sexo não é determinada por cromossomos, mas pela
ação diferencial dos genes. A progênie, em relação ao sexo, é formada segundo as
proporções mendelianas. Um exemplo refere-se a ação
dos genes Ba/ba e Ts/ts no milho.
No
milho, esses genes atuam do seguinte maneira:
Ba - :
planta normal
ba
ba : planta com talo estaminado, mas sem espiga
Ts - : planta normal
ts
ts : planta com pendão substituído por uma estrutura
pistilada
Do cruzamento entre plantas duplo-heterozigotas (Ba ba Ts ts)
surge na descendência:
9/16 de
plantas normais (Ba_ Ts_)
3/16 de
plantas unissexuais masculinas (ba
ba Ts_)
3/16 de
plantas unissexuais femininas (Ba_ ts ts)
1/16 de
planta unissexual feminina pouco produtiva (ba ba ts
ts)
![]() Volta
Volta
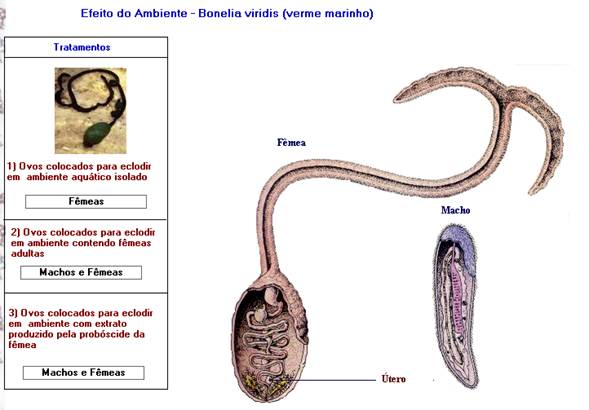
EFEITO DO AMBIENTE
Neste caso, machos e fêmeas têm a mesma constituição genética, mas a
diferenciação dos órgãos sexuais é dependente de estímulos ambientais. Um exemplo
de determinação do sexo através do ambiente é encontrado no verme marinho Bonellia viridis,
em que o macho, bem menor que a fêmea, vive dentro do aparelho reprodutivo das
fêmeas. A fêmea libera os ovos, já fertilizados, no meio aquático, os quais, depois
de eclodidos, diferenciam-se em machos e fêmeas. Para sobreviver, os machos
penetram através da abertura superior da fêmea e atingem o ovário dela.
As evidências de que a determinação do sexo é feita por estímulo do ambiente
surgiram no seguinte experimento:
Tratamento 1: Ovo colocado para eclodir em um ambiente
aquático isolado. Neste caso, todos os ovos eclodiram, dando origem a apenas
fêmeas.
Tratamento
2: Ovo colocado para eclodir em ambientes contendo
fêmeas adultas. Os ovos eclodiram, dando origem a machos e fêmeas. Os machos
migraram para o interior das fêmeas adultas.
Tratamento 3: Ovos colocados para eclodir em ambiente
que continha extrato de fêmeas. Os ovos eclodiram, dando origem a machos e
fêmeas. Os machos não sobreviveram pela ausência de fêmeas adultas.
Neste experimento ficou demonstrado que as fêmeas surgem em qualquer situação,
mas, para a diferenciação do sexo masculino, é necessária a presença da fêmea
ou de substâncias produzidas por elas.
![]() Volta
Volta
ABERRAÇÕES SEXUAIS NA ESPÉCIE HUMANA
Além do padrão normal 2A +XY e 2A + XX (A refere-se a um conjunto autossomal com 22 cromossomos), existem, na população
humana, indivíduos com excesso ou falta de cromossomos sexuais, classificados
como portadores de aberrações sexuais. Podem ser citados os seguintes exemplos:
Síndrome de Klinefelter
Trata-se da reversão sexual do indivíduo masculino em direção ao sexo feminino.
Ocorre com uma taxa de 2 a 3 em cada 1.000 indivíduos. Os sintomas graves são:
porte alto, tendência a feminilização, seios grandes
e retardamento mental. Entretanto, alguns são imperceptíveis. Esses indivíduos
podem se casar e a consumação pode ser efetuada de forma legal. A Constituição
cromossômica é de 44 autossomais XXY. É considerado também o indivíduo XXYY.
Indivíduos XXXY ou XXXXY são considerados como extremo de Klinefelter.
A causa principal é a anomalia meiótica, por exemplo, uma não-disjunção
das cromátides tanto na ovogênese quanto na
espermatogênese (em menor probabilidade).
Síndrome de Turner
Reversão sexual do indivíduo do sexo feminino em direção ao sexo masculino.
Ocorre com uma taxa de 0,2 a 0,3 em cada 1.000 indivíduos. Como sintomas mais
característicos destacam-se a estatura mais baixa, o pescoço alado, e a
subfertilidade. A constituição cromossômica é 44 autossomais + XO.
Síndrome XYY
São indivíduos agressivos e de pouca inteligência. São comumente encontrados em
hospícios e hospitais, entretanto a razão de serem encontrados também em
prisões não está associada a taras sexuais.
![]() Volta
Volta
CROMATINA SEXUAL
Em adição às várias distinções entre os sexos com relação aos cromossomos
sexuais (tamanho, propriedades tinturiais, ativação
gênica etc.), existe também uma importante diferença no que diz respeito a coloração de núcleos somáticos.
De
acordo com Barr e Bertran,
nas fêmeas de mamíferos, existe um local de coloração no núcleo que não é
encontrado nos machos. Esses sítios coloridos são denominados "corpúsculo de Barr", "corpúsculo de cromatina positiva" ou
"cromatina sexual". O número de corpúsculos de Barr
constatado em células somáticas, em alguns indivíduos, é:
|
Indivíduo |
No. de
corpúsculos |
|
Homem normal (XY) |
0 |
|
Mulher normal (XX) |
1 |
|
Síndrome de Klinefelter
(XXY) |
1 |
|
Síndrome de Turner (XO) |
0 |
|
Extremo de Klinefelter
(XXXY) |
2 |
|
Síndrome XYY |
0 |
Com as observações anteriores, ficou constatado que:
Nº de corpúsculo = Nº de cromossomo X - 1
De acordo com a hipótese apresentada por Lyon e Russel, os corpúsculos seriam o
resultado da inativação, compactação e heterocromatização de todos, exceto um,
cromossomo X do indivíduo. Fêmeas de mamíferos (XX) devem apresentar um destes
cromossomos inativo, de maneira que o relacionamento de dosagem entre
cromossomos sexuais eucromáticos e cromossomos autossomais seria o mesmo que
nos tecidos somáticos dos machos.
A heterocromatização é facultativa, ou seja, ocorre apenas em determinados
estágios da célula. Assim, uma mulher de constituição cromossômica XX é capaz
de formar óvulos contendo cromossomo X ativo.
A
inativação é casual. Uma mulher XmXp,
em qu Xm é o
cromossomo X de origem materna e Xp é o
cromossomo X de origem paterna pode apresentar, dependendo da célula somática,
um ou o outro cromossomo X inativo.
Na espécie humana, a heterocromatização só é observada após o 16º dia de
gestação. Esse pequeno período de ativação do cromossomo X é suficiente para
causar as variações encontradas entre um indivíduo XY e XXY.
Um exemplo da inativação casual do cromossomo X é encontrado em gatos. Nesses
animais, o padrão de pelagem preto e amarelo é condicionado por um gene ligado
ao sexo, e o padrão branco, por outro gene autossomal.
As seguintes cores são verificadas:
|
Fêmeas |
Machos
|
|
XAXA preta (preta e branca) |
XAY preto (preto e branco) |
|
XAXB preta e amarela (preta, amarela e branca) |
XBY amarelo (amarelo e branco) |
|
XBXB Amarela (amarela e
branca) |
![]() Volta
Volta
GINANDROMORFISMO
Ginandromorfos são mosaicos sexuais nos quais algumas
partes do animal apresentam características femininas e outras masculinas. Em
alguns casos, tanto as genitálias e gônodas
masculinas quanto as femininas podem estar presentes no mesmo animal. A freqüência natural tem sido de ½.000
ou 1/3.000 moscas. Entretanto, muitos desses ginandromorfos
apresentam poucas secções revertidas. Os ginandromorfos
bilaterais são mais raros. A freqüência pode ser
aumentada quando o indivíduo apresenta um cromossomo X em anel (ligado pelas extremidades).
Os ginandromorfos bilaterais têm sido explicados como uma
irregularidade na mitose da primeira clivagem. Um exemplo interessante foi
observado na descendência de cruzamentos envolvendo machos de olho barra e
fêmea normal. O caráter forma do olho é regulado por um gene ligado ao sexo em
que o "alelo" B determina o olho barra e o alelo recessivo b, o olho
normal.
Foi considerado o seguinte cruzamento: fêmea normal (XbXb)
e macho barra (XBY). A descendência feminina foi do tipo XBXb. Uma
anomalia no processo fez com que a célula zigótica
originasse células com constituição cromossômica diferente em relação aos
cromossomos sexuais. Foram formadas células XBXb e XbO,
de forma que o indivíduo adulto era do tipo XBXb
/ XbO Assim, os tecidos e órgãos que
tiveram origem de XBXb
manifestavam características sexuais femininas, e aqueles com origem em XbO, manifestavam características masculinas.
Além das diferenças proporcionadas pelo padrão sexual, observa-se no ginandromorfo bilateral, originado do referido cruzamento,
um olho barra (XBXb)
e outro normal (XbO)
Um ginandromorfo se diferencia de um intersexo quanto
à origem e à constituição cromossômica. O ginandromorfo
tem origem numa irregularidade mitótica, enquanto o intersexo é formado por uma combinação
gamética que resulta num índice sexual entre 0,5 e 1,0. O ginandromorfo
apresenta, em
um mesmo indivíduo, células com diferentes números de cromossomos. No
intersexo, o número de cromossomos é constante para todas as células do
indivíduo.